Упрощенная HTML-версия


лирования реализуются пути, описанные для случаев без корреляций. Это
может быть объяснено нарушением схемы корреляций нуклеотидных за
мен и, соответственно, уменьшением
значения
приспособленности и пол
ной элиминацией мутировавших последовательностей и их потомков, да
же при высоком уровне гомологии внутри группы. Тогда место группы
наиболее приспособленных последовательностей занимает другая субпо
пуляция с достаточно большим значением приспособленности у каждой
последовательности. Такой режим быстрого восстановления при режиме
хаотических колебаний численности группы наиболее приспособленных
последовательностей возможен при наличии высокого уровня гомологии
(> 94%) внутри всей популяции.
Наличие корреляций в процессе моделирования ведет также к сни
жению значения максимальной достигаемой приспособленности в попу
ляции и большему генетическому разнообразию последовательностей, по
лученных в результате моделирования.
-
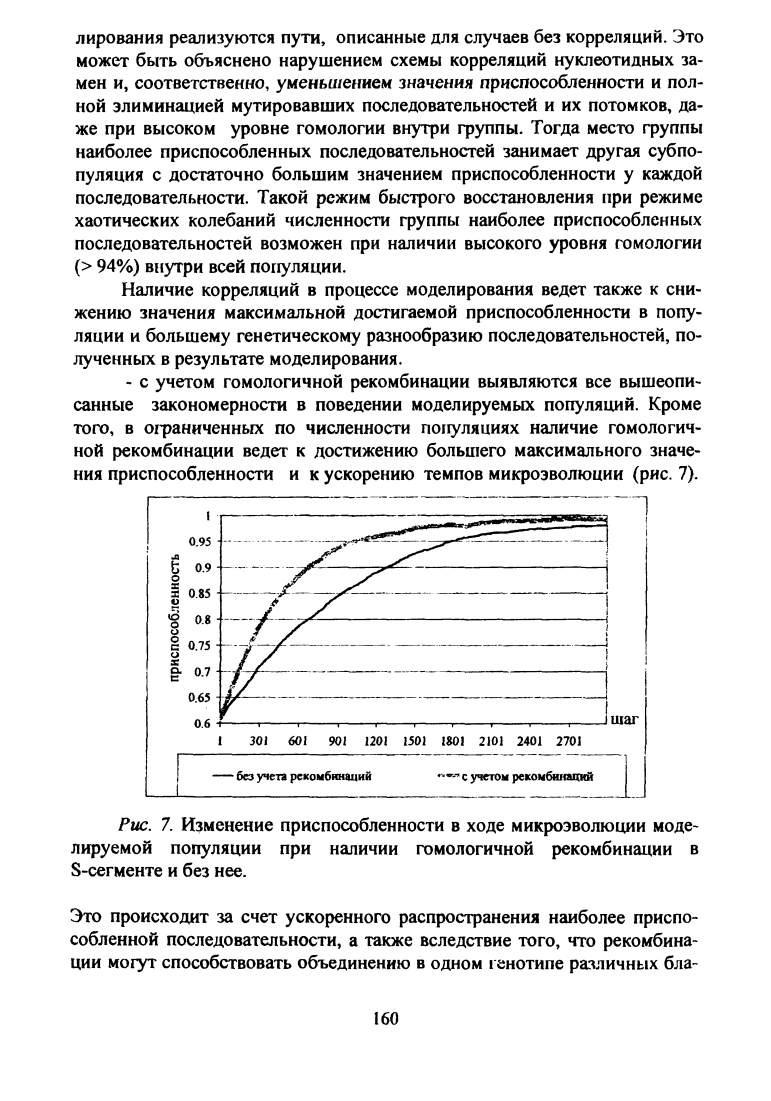
с учетом гомологичной рекомбинации выявляются все вышеопи
санные закономерности в поведении моделируемых популяций. Кроме
того, в ограниченных по численности популяциях наличие гомологич
ной рекомбинации ведет к достижению большего максимального значе
ния приспособленности и к ускорению темпов микроэволюции (рис. 7).
Рис
. 7. Изменение приспособленности в ходе микроэволюции моде
лируемой популяции при наличии гомологичной рекомбинации в
S-сегменте и без нее.
Это происходит за счет ускоренного распространения наиболее приспо
собленной последовательности, а также вследствие того, что рекомбина
ции могут способствовать объединению в одном генотипе различных бла
160